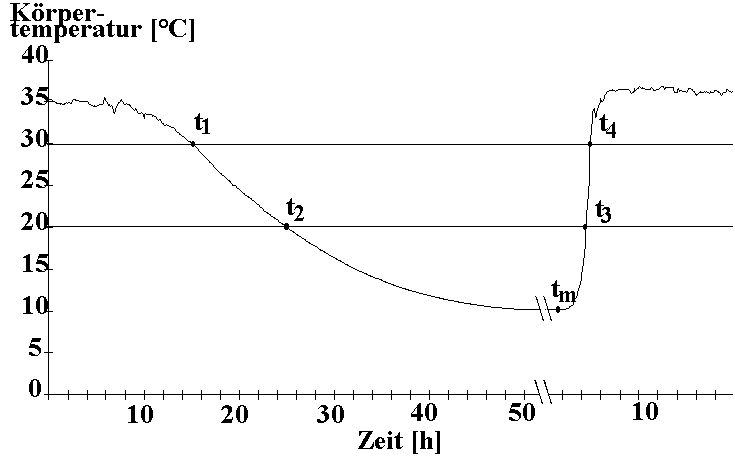
Abb. 1: Ablauf eines typischen Winterschlaf-schubs in einer zeitlichen Auflösung von 0,5min und einer Meßgenauigkeit von 0,1°C mit den Zeitpunkten: t1 (Beginn, Tb < 30°C); t2 (Tb < 20°C); tm (Tb=Tmin); t3 (Tb > 20°C) und t4 (Ende, Tb > 30°C).
Der Winterschlaf des Europäischen Feldhamsters (Cricetus cricetus) unter kontrollierten Belichtungs- und Temperaturbedingungen
Hibernation of the European hamster (Cricetus cricetus) under controlled photoperiod and ambient temperature.
T. Waßmer & F. Wollnik, Fakultät für Biologie, Universität Konstanz, Postfach 5560, M618, D-78434 Konstanz
Das Winterschlafverhalten des Europäischen Feldhamsters (Cricetus cricetus) wurde durch Langzeitmessungen der Körpertemperatur (Tb) mittels in die Bauchhöhle implantierter Telemetriesender (1,5 x 2,5 cm; 6 g) untersucht. Hierzu wurden vier erwachsene Feldhamstermännchen (ca. 1,5 Jahre alt) im Oktober 1992 aus dem Freiland in eine Klimakammer überführt. Dort wurden die Tiere einzeln in jeweils ca. 1m3 großen nach oben durch ein Drahtgitter verschlossenen PVC Boxen gehalten, die mehrere Wohnkammern aus PVC enthielten und bis ca. 0,2m mit Erde gefüllt waren. Wasser und pelletierte Standarddiät wurde ad libitum gereicht. Als Vorbereitung auf den Winterschlaf wurden die Tiere für 4 Wochen unter Kurztagbedingungen (LD8:16) gehalten und anschließend die Temperatur schrittweise auf 8 ± 1°C abgekühlt.
| Ein typischer Winterschlafschub (WS, Abb.1) dauerte zwischen 15,3h und 82,4h (M±SD=48,0±16,8h; N=36; definiert als t1-t4) und ereichte in der linearen Abkühlphase (t1-t2) eine Abkühlrate von 1,2±0,2°C/h. Die minimale Tb (Tmin = 10,5±2,0°C) wurde meist kurz vor Beginn der Aufwärmphase erreicht und war signifikant mit der Dauer des Torpors korreliert (rs=-0,79; p<0,001; t-Test; N=46). Die lineare Aufwärmrate (t3-t4) betrug 14,7±4,5°C/h. Neben den typischen WS wurden sogenannte "shallow torpors bouts" (ST) beobachtet, die mit einer mittleren Dauer von 4,1±2,6h (N=10) wesentlich kürzer waren und sich durch eine geringere Temperaturabsenkung (20°C<Tmin <30°C) und geringere Aufwärmraten (8,5±2,8°C/h; N=8) von den WS unterschieden (Kruskal Wallis ANOVA, p<0,001, N=44). |
Abb. 1: Ablauf eines typischen Winterschlaf-schubs in einer zeitlichen Auflösung von 0,5min und einer Meßgenauigkeit von 0,1°C mit den Zeitpunkten: t1 (Beginn, Tb < 30°C); t2 (Tb < 20°C); tm (Tb=Tmin); t3 (Tb > 20°C) und t4 (Ende, Tb > 30°C). |
Es wurden beachtliche interindividuelle Unterschiede im Hinblick auf die Anzahl der WS und ST beobachtet. Zum anderen unterschieden sich die Tiere im Hinblick auf die Dauer (p£ 0,01; N=46) und die Abkühlrate (p<0,05; N=44) der WS (Kruskal Wallis ANOVA). Der Beginn (t1) der einzelnen WS zeigte bei nur einem Tier einen statistisch signifikanten Mittelvektor bei 23 Uhr 12 min MEZ (r=0,67; p£ 0,001; Rayleigh-Test; N=14). Ebenfalls fand sich nur bei einem Tier ein statistisch signifikanter Mittelvektor für das Ende der WS (9 Uhr 4 min MEZ; r=0,63; p£ 0,03; Rayleigh-Test; N=9).
Die vorliegenden Ergebnisse unterstützen die Auffassung, daß das Winterschlafverhalten des Europäischen Feldhamsters nicht nur durch exogene Faktoren (Umgebungstemperatur, Photoperiode) bestimmt wird, sondern vielmehr durch endogene Faktoren (Zusammenhang zwischen der Länge der WS und Tmin, eventuell circadiane Rhythmik) beeinflußt wird (Michener GR 1992: Oecol 89,397; Schmidt B, Wollnik F 1993: Verh. Dtsch Zool Ges 86.1, 282).
Gefördert durch die DFG (WO 354/5-1).
![]() -
BACK TO PUBLICATIONS - ZURÜCK
ZUR PUBLIKATIONSLISTE
-
BACK TO PUBLICATIONS - ZURÜCK
ZUR PUBLIKATIONSLISTE